
Yellow-bellied Marmot
Distribution, Abundance, and Seasonality
The yellow-bellied marmot is common and widespread in, or near, rocky areas in the Sierra Nevada and Cascades from southern Tulare Co. north to the Oregon border, and in the White Mts. of California and Nevada. Optimum habitats are alpine dwarf-shrub, perennial grassland, wet meadow, subalpine conifer, and open stands of lodgepole pine forest. Also found in open, grassy understories of mixed conifer, montane riparian, red fir, eastside pine, montane chaparral, sagebrush, bitterbrush, low sage, aspen, Jeffrey pine, and pinyon-juniper habitats.
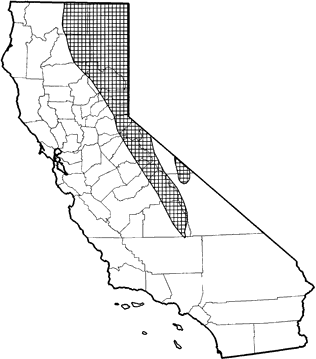
Range Map
Specific Habitat Requirements
Feeding: This herbivore feeds on seeds, flowers, leaves, and stems of a wide variety of grasses, forbs, and shrubs, and it occasionally eats insects. It forages on the ground. Fat (to 50% of body weight) is accumulated prior to hibernation. Marmots appear to avoid plants with known high concentrations of secondary compounds such as alkaloids or tannins (Armitage 1979).
Cover: Uses rocky areas and talus for cover. Digs a burrow system as a refuge and hibernaculum. Burrow usually is under rocks, but also may be under tree roots or buildings.
Reproduction: Builds a nest of dry grasses in the burrow system.
Water: Drinks water while active. Captive animals from a xeric habitat drank less and produced less, and more concentrated, urine than animals from a mesic habitat (Ward and Armitage 1981b). In the xeric habitat, 73-74% of the total water intake came from drinking, and the remainder came from vegetation and metabolic water.
Pattern: Prefers rocky outcrops and talus slopes with nearby grasses and forbs. Uses rocks for shelter, sunning, and observing. Uses open stages of forests if an understory of grasses and forbs is present. Requires water source. May avoid areas of heavy grazing. In the Sierra Nevada, large populations may be found in, and around, meadows. Small "satellite" populations may occur in less preferred sites. These populations have lower reproductive rates and high turnover (Frase and Hoffmann 1980).
Species Life History
Activity Patterns: Diurnal; hibernates from September or October to April or May. Some low-elevation populations begin hibernation/aestivation in June or July. These populations may emerge in March. Brief emergences may occur before the final emergence. Males are the first to hibernate, followed by females and young, which remain active to gain sufficient fat for hibernation. Daily activity mostly is bimodal. After a period of sunning, there is a mid-morning peak of foraging activty followed by another peak in late afternoon. This pattern is subject to seasonal variation. Along the Columbia River a population is becoming somewhat nocturnal because of human hunting pressure and disturbance (Ingles 1965). Circannual rhythms of food consumption and metabolic rate persist in the laboratory; two populations held unique rhythms related to their habitat (Ward and Armitage 1981a).
Seasonal Movements / Migration: None.
Home Range: Home ranges in Colorado varied from 2.2-10 ha (4.9-24.7 ac) (Armitage 1974). Most marmots are found in loose colonies (Barash 1974, Svendsen 1974), though some occur in pairs, or alone.
Territory: Marmot colonies consist of one, or more, harems, each harem defended by a territorial male. In Colorado, male territories ranged from 0.2-2.0 ha (0.5-4.9 ac), with a mean of 0.54-0.77 ha (1.2-1.7 ac) (Armitage 1974). Scent marking is an expression of dominance (Armitage 1976).
Reproduction: Mates in the first 2 wk after emergence. After a 30-day gestation, young born March to June, depending on elevation. The single litter usually is 4-6 young, ranging from 3-8. Lactation lasts 20-35 days. Those young that wean early have more time to accumulate fat for hibernation, and a higher probability of survival through the winter. All males, and many females, disperse as yearlings at low and middle elevations, or as 2-yr-olds at high elevations. Yearling females rarely reproduce. Only 25% of 2-yr-old females reproduced at 2900 m (9300 ft) (Armitage and Downhower 1974), while at 1700-2200 m (5600-7000 ft) 40% of 2-yr-old females reproduced. At high elevations, females may reproduce in alternate years. The amount and quality of spring food, and availability of spring hibernacula, influence the reproductive success of populations.
Niche: This hibernating herbivore occurs with several other potentially competing herbivores: the pika, Belding's ground squirrel, and various species of microtus. Preyed upon by badgers, coyotes, eagles, owls, and wolverines. Host to a tick that may carry Rocky Mountain spotted fever.
Sources & References
California Department of Fish and Game, 1999.
California's Wildlife, Sacramento, CA.
Written by: V. Johnson, J. Harris, reviewed by: H. Shellhammer, edited by: R. Duke, S. Granholm
Anderson, D. C., K. B. Armitage, and R. S. Hoffmann. 1976. Socioecology of marmots: female reproductive strategies. Ecology 57:552-560. Armitage, K. B. 1962. Social behavior of a colony of the yellow-bellied marmot (Marmota flaviventris). Anim. Behav. 10:319-331. Armitage, K. B. 1974. Male behavior and territoriality in the yellow-bellied marmot. J. Zool., London 172:233-265. Armitage, K. B. 1976. Scent marking by yellow-bellied marmots (Marmota flaviventris). J. Mammal. 57:583-584. Armitage, K. B. 1977. Social variety in the yellow-bellied marmot: a population-behavioral system. Anim. Behav. 25:585-593. Armitage, K. B. 1979. Food selection by yellow-bellied marmots. J. Mammal. 60:628-629. Armitage, K. B., and J. F. Downhower. 1974. Demography of yellow-bellied marmot populations. Ecology 55:1233-1245. Barash, D. P. 1974. The evolution of marmot societies: a general theory. Science 185:415-420. Downhower, J. F., and K. B. Armitage. 1981. Dispersal of yearling yellow-bellied marmots (Marmota flaviventris). Anim. Behav. 29:1064-1069. Frase, B. A., and R. S. Hoffmann. 1980. Marmota flaviventris. Mammal. Species No.135. 8pp. Hock, R. J. 1969. Thermoregulatory variations of high-altitude hibernators in relation to ambient temperature, season, and hibernation. Fed. Procs. 28:1047-1052. Ingles, L. G. 1965. Mammals of the Pacific states. Stanford Univ. Press, Stanford, CA. 506pp. Kilgore, D. L., Jr., and K. B. Armitage. 1978. Energetics of a yellow-bellied marmot population. Ecology 59:78-88. Nee, J. A. 1969. Reproduction in a population of yellow-bellied marmots (Marmota flaviventris). J. Mammal. 50:756-765. Svendsen, G. E. 1974. Behavioral and environmental factors in the spatial distribution and population dynamics of a yellow-bellied marmot population. Ecology 55:760-771. Ward, J. M., and K. B. Armitage. 1981a. Circannual rhythms of food consumption, body mass and metabolism in yellow-bellied marmots (Marmota flaviventris). Comp. Biochem. Physiol. A69:621-626. Ward, J. M., and K. B. Armitage. 1981b. Water budgets of montane-mesic and lowland-xeric populations of yellow-bellied marmots (Marmota flaviventris). Comp. Biochem. Physiol. A69:627-630. Webb, D. R. 1980. Environmental harshness, heat stress, and Marmota flaviventris. Oecologia (Berlin) 44:390-395.
California Animal Facts | California's Wildlife